Die Evolution der sexuellen Kommunikation von Motten
Forschungsbericht (importiert) 2013 - Max-Planck-Institut für chemische Ökologie
Mottenweibchen produzieren artspezifische Pheromone und locken damit Männchen an. Kleinste Änderungen des Dufts sowie die Neigung von Männchen, einem veränderten Geruch zu folgen, führt zu vermindertem Paarungserfolg und sollte daher negativ selektiert werden. Somit stehen wir vor einem Rätsel: Wie konnten sich trotz starker Selektion unterschiedliche Lockstoffe bei verschiedenen Mottenarten entwickeln? Neue Studien geben Aufschluss: Genetische Variation kann sogar innerhalb einer Art bestehen. Zum Teil ist diese auch für die Evolution von Unterschieden zwischen Arten verantwortlich.
Sexuelle Kommunikation bei Motten
Die Falter der meisten Mottenarten haben eine kurze Lebenszeit und sind nur in der Nacht aktiv. Damit sich die Männchen und Weibchen einer Art fortpflanzen können, müssen sie sich zur Paarung finden. Weibchen bleiben ortsgebunden und machen die Männchen durch Abgabe eines Sexuallockstoffs auf sich aufmerksam: eine Duftmischung, die in der Regel aus zwei oder mehr flüchtigen Substanzen besteht. Die Männchen reagieren hochsensibel auf das weibliche Pheromon und spüren die Duftquelle auf; Balz und Paarung folgen. Es existieren Tausende verschiedener Pheromonmischungen; jede Art produziert eine ganz bestimmte Duftkomposition und reagiert auch nur auf diese. Diese Spezifität stellt sicher, dass Paarungen zwischen Weibchen und Männchen verschiedener Arten verhindert werden.
Die Weibchen produzieren die einzigartige Duftmischung mithilfe einer „Chemiefabrik“, die sich in einer speziellen Drüse an der Spitze ihres Hinterleibs befindet. Sie enthält Enzyme, die verantwortlich für die Biosynthese der unterschiedlichen Komponenten des Lockstoffs sind. Die Enzyme, darunter Fettsäure-Synthasen, Desaturasen, Reduktasen und Acetyltransferasen, arbeiten zusammen, damit das optimale Verhältnis von größeren und kleineren Bestandteilen der Pheromonmischung gebildet wird. Die Pheromone werden in die nächtliche Luft freigesetzt, wenn das Weibchen die Oberfläche der Drüse entblößt. Der kontinuierliche Strom der flüchtigen Substanzen bildet eine Duftfahne, die vom Wind über große Distanzen getragen wird. So entsteht die Duftspur, der die Männchen folgen können.
Das Männchen entdeckt das chemische Signal mithilfe seiner außergewöhnlich sensiblen Antennen, die bereits wenige Pheromonmoleküle aufspüren können. Spezielle Neuronen in den winzigen Härchen auf der Antenne tragen Rezeptor-Proteine, an die sich die Duftstoffe spezifisch binden. Verschiedene Arten haben unterschiedliche Rezeptoren, die jeweils ein ganz bestimmtes Pheromon binden. Die Bindung aktiviert die Neuronen. Nachdem das Signal an die darauf spezialisierten Bereiche im Gehirn weitergeleitet wurde, ist die Motte stimuliert, gegen den Wind und in Richtung der Duftquelle zu fliegen. Sobald ein Bestandteil der Pheromonmischung fehlt oder ein ungewollter Bestandteil hinzugekommen ist, wird das Flugverhalten unterbunden. Das Männchen fliegt weiter gegen den Wind, solange es auf die richtige Duftmischung stößt. Verliert das Mottenmännchen die Duftfahne, ändert es die Richtung im Zickzackflug, bis es die Duftfahne wiedergefunden hat. Dieses Verhalten führt es letztendlich zum Weibchen.
Das feinabgestimmte und spezifische Kommunikationssystem ist ein Wunder der Evolution. Dennoch stellt es ein Dilemma dar, die vielen Tausend unterschiedlichen Pheromonmischungen zu erklären, die sich im Laufe der Evolution entwickelt haben. Lange Zeit haben Pheromon-Forscher angenommen, dass das Kommunikationssystem jeder einzelnen Art durch eine starke stabilisierende Selektion aufrechterhalten wurde. Produziert ein Weibchen einen Lockstoff, der von dem optimal anlockenden Sexualpheromon abweicht, wird es von den Männchen seiner Art weniger wahrscheinlich gefunden und der Fortpflanzungserfolg wird geringer sein. Folgt das Männchen einem Pheromon, das ebenfalls von der optimalen Duftmischung abweicht, wird es womöglich von Weibchen einer anderen Art irregeführt und der Paarungserfolg ist ebenfalls geringer. Schwierig wird es jedoch, wenn man die Entstehung einer neuen Art mit einem veränderten Pheromonsystem erklären möchte, denn dafür wäre eine bereits bestehende Variation innerhalb der ursprünglichen Art erforderlich gewesen und die hätte eigentlich negativ selektiert werden müssen.
Die Lösung dieses Dilemmas erschließt sich nur, wenn man diesen Prozess auf genetischer Ebene versteht. Derzeit ist das schwierig, denn die meisten Gene, die der weiblichen Biosynthese und der männlichen Wahrnehmung der Sexualpheromone zugrundeliegen, sind noch nicht bekannt. In jüngerer Zeit gab es einige Untersuchungen zur Identifizierung dieser Gene. Sie haben wichtige Informationen geliefert, um zu verstehen, wie sich Pheromonsysteme im Laufe der Evolution verändern konnten. Im Folgenden werden Ergebnisse über die Entdeckung von Genen, die in der weiblichen Pheromondrüse exprimiert werden, dargestellt und genetische Studien zusammengefasst, in denen die genetische Variation innerhalb von Arten sowie zwischen nahe verwandten Arten identifiziert wird, die sich auf die Pheromonproduktion auswirkt.
Genetische Variationen der Pheromonproduktion innerhalb einer weit verbreiteten Art
Die Amerikanische Tabakeule Heliothis virescens ist eine weit verbreitete nachtaktive Motte, die in Nord- und Südamerika vorkommt (Abb. 1). Ihre Raupen sind Generalisten, die sich von 37 Pflanzenarten aus 14 verschiedenen Pflanzenfamilien ernähren, darunter viele Nutzpflanzen, wie z.B. Tabak, Baumwolle, Soja, Tomate, Sonnenblume und Kichererbse. Sie ist daher ein bedeutender Ernteschädling.
 und Motte (rechts) der Amerikanischen Tabakeule Heliothis virescens")

Abb. 1: Raupe (links) und Motte (rechts) der Amerikanischen Tabakeule Heliothis virescens
Die Pheromonzusammensetzung dieser Art wurde ursprünglich in der Hoffnung analysiert, dass sie in Form einer Pheromonfalle zur Schädlingsbekämpfung eingesetzt werden könnte. Die meisten der frühen Untersuchungen aus den 1980er Jahren beschränkten sich darauf, die optimale Duftmischung zu bestimmen, und beachteten die Variabilität zwischen den verschiedenen Weibchen überhaupt nicht. Allerdings erbrachte eine 2005 begonnene Langzeitstudie mit Stichproben, dass diese Variabilität beträchtlich ist [1]. Einige Weibchen produzierten einen deutlich geringeren Anteil von (Z)-11-Hexadecenal (Z11-16:Ald), einem Bestandteil des Pheromons, der beim Anlocken der Männchen eine wesentliche Rolle spielt, und einen deutlich höheren Anteil an Hexadecanal (16:Ald), der wiederum sehr wenig zur Attraktivität bei den Männchen beiträgt. Beide Verbindungen sind in ihrer chemischen Struktur sehr ähnlich und unterscheiden sich lediglich in einer Doppelbindung der Kohlenwasserstoffkette (Abb. 2). Deshalb wurde bei diesen Weibchen die niedrigere Aktivität eines Desaturase-Enzyms vermutet, das Doppelbindungen einführt. Warum jedoch diese Weibchen eine suboptimale Duftmischung produzierten, blieb rätselhaft.
 und Heliothis subflexa (Hs). Für jede Einzelkomponente ist ihr prozentualer Anteil im Pheromon dieser Art angegeben. Bestandteile, die wesentlich der Anlockung von Männchen dienen, sind grün dargestellt.")
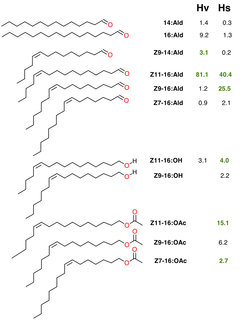
Abb. 2: Bestandteile des Pheromons der Weibchen von Heliothis virescens (Hv) und Heliothis subflexa (Hs). Für jede Einzelkomponente ist ihr prozentualer Anteil im Pheromon dieser Art angegeben. Bestandteile, die wesentlich der Anlockung von Männchen dienen, sind grün dargestellt.
Um die genetischen Grundlagen dieses Unterschiedes zu erforschen, wurden zwei Populationen im Labor entwickelt: Eine der Populationen wurde für hohe Mengen von 16:Ald selektiert (hohe Linie), die andere für niedrige Mengen (niedrige Linie). Beide Populationen wurden miteinander gekreuzt und die hybriden Nachkommen zur niedrigen Linie zurückgekreuzt. Pheromone aus den Drüsen der Weibchen der zurückgekreuzten Generation wurden extrahiert und mithilfe genetischer Marker auf allen 31 Chromosomen dieser Art verglichen. Eine Variation auf Chromosom 4 korrelierte signifikant mit der Menge an 16:Ald, was darauf hindeutete, dass ein oder mehrere Gene auf diesem Chromosom den Unterschied zwischen Weibchen der hohen und der niedrigen Linie verursacht [1].
Die Genexpression mehrerer Enzyme im Biosyntheseweg des Pheromons wurde gemessen und dabei wurde ein spezifisches Desaturase-Enzym gefunden, das in Weibchen, die weniger Z11-16:Ald und mehr 16:Ald produzierten, schwächer exprimiert wurde. Obwohl die niedrigere Desaturase-Aktivität die geringere Produktion von Z11-16:Ald erklären könnte, kommt das Gen für dieses Enzym nicht auf Chromosom 4 vor. Stattdessen scheint die Expression dieser Desaturase von einem transkriptionellen Repressor gesteuert zu werden, dessen Gen auf Chromosom 4 zu finden ist. Ein Aktionsmodell dieses Repressors, das zu allen bekannten Daten passt, trifft eine ungewöhnliche Vorhersage: Die für das Repressor-Gen heterozygoten Tiere sollten mehr Z11-16:Ald produzieren als die homozygoten, denn es ist ja nur die Hälfte des Repressors vorhanden. Dies würde einen ausbalancierten Polymorphismus herbeiführen − eine Art natürlicher Selektion, die beispielsweise auch den Polymorphismus für Sichelzellenanämie − einer erblichen Erkrankung der roten Blutkörperchen − in menschlichen Populationen aufrechterhält: Bei Bewohnern afrikanischer Malariaregionen treten gesunde Überträger des Sichelzellenallels kontinuierlich auf, denn: Sie sind gegen Malaria resistent und haben deshalb einen evolutionären „Heterozygotenvorteil“, obwohl viele ihrer roten Blutkörperchen gar nicht intakt sind. Das aber hat den positiven Nebeneffekt, dass sich der Malariaerreger wegen Mangel an intakten roten Blutkörperchen nicht intensiv vermehren kann. Damit vergleichbar erhält sich so also auch die genetische Variabilität innerhalb der Mottenweibchen aufrecht, weil die Mottenmännchen hochsensibel auf die optimale Mischung mit dem höchsten Anteil von Z11-16:Ald reagieren [1].
Ähnliche genetische Variation innerhalb und zwischen den Arten
Eine nahe Verwandte der Tabakeule ist die Motte Heliothis subflexa, die im Gegensatz zu Heliothis virescens ein Wirtsspezialist ist und nur Physalis-Früchte verspeist. Dazu gehören die süßen, gelb-orangen, beerenähnlichen Früchte, die in Europa beliebte Salatzutaten sind, aber auch die sauren, grünen Tomatillos, die wir von mexikanischen Salsa-Gerichten kennen. H. subflexa kommt zahlreich im westlichen Mexiko vor, wo Tomatillos angebaut werden, und kann dort zu einem bedeutenden Schädling dieser Nutzpflanzen werden. Weniger Exemplare dieser Art finden sich im Südosten der USA, wie z. B. in North Carolina, wo die einzigen Physalis-Arten als unkultivierte Kräuter vorkommen und H. virescens häufiger anzutreffen ist. Ein interessantes Muster der Pheromonvariation in H. subflexa ist im Hinblick auf die geografische Verbreitung erkennbar: Essigsäureester wie Z11-16:OAc (Abb. 2) kommen in höherer Konzentration im Pheromon von H. subflexa-Weibchen in North Carolina vor, nehmen dann im geografischen Verlauf kontinuierlich ab und sind im Westen von Mexiko nur noch in niedrigen Konzentrationen nachweisbar [2].
Essigsäureester sind wesentliche Bestandteile der Pheromonzusammensetzung von H. subflexa: Sie locken Männchen dieser Art an, wirken jedoch auf Männchen der verwandten Art H. virescens abschreckend. Tatsächlich produzieren die Weibchen von H. virescens überhaupt keine Essigsäureester. Allerdings sind die Pheromone beider Arten ansonsten sehr ähnlich. Es konnte gezeigt werden, dass H. subflexa-Weibchen, die derart verändert wurden, dass sie nur noch sehr geringe Mengen Essigsäureester produzieren konnten, auch einige H. virescens-Männchen anlockten und insgesamt einen geringeren Reproduktionserfolg aufwiesen [3]. Hohe Konzentrationen von Essigsäureestern wirken bei Männchen der Art H. virescens als STOPP-Signal, was in North Carolina, wo diese Männchen viel häufiger vorkommen, wesentlich wichtiger ist als im Westen von Mexiko.
Um die genetische Grundlage der Essigsäureester-Variation in H. subflexa zu untersuchen, wurden Populationen aus dem westlichen Mexiko, die niedrige Mengen der Verbindung in ihrem Pheromon produzierten, mit Populationen aus North Carolina, die hohe Essigsäureester-Konzentrationen in ihrem Pheromon aufwiesen, miteinander gekreuzt. Die genetische Variation für Marker auf allen 31 Chromosomen wurde in Weibchen der zurückgekreuzten Generation untersucht. Eine ähnliche Rückkreuzung zwischen H. subflexa und H. virescens wurde ebenfalls analysiert, um zu untersuchen, warum H. virescens keine Essigsäureester produziert. Es wurden zwei unterschiedliche Chromosomen von H. virescens identifiziert, die Gene enthalten, welche die Produktion von Essigsäureester unterdrücken. Eines dieser Chromosomen kontrolliert auch die Variation der Essigsäureester-Konzentrationen zwischen den östlichen und westlichen Populationen von H. subflexa. Dies ist der erste Hinweis darauf, dass die genetische Variabilität innerhalb einer Art auch eine Rolle spielen könnte, wenn es um die Aufrechterhaltung von Kreuzungsbarrieren zwischen verschiedenen Arten geht. Darüber hinaus ergibt sich, wie der erste Schritt hin zur Entstehung einer neuen Pheromonmischung aussehen könnte [4].
Ausblick
Die genetischen Untersuchungen wurden durch neueste DNA-Sequenzierungsmethoden (next-generation sequencing) ermöglicht. Weitere Fortschritte bei der Identifizierung der Gene werden durch die vollständige Genom-Sequenzierung beider Arten unterstützt. Die Ergebnisse verweisen auf die Bedeutung des genetischen Ansatzes, der bislang in der Pheromonforschung meist vernachlässigt wurde, und auf den Wert detaillierter Untersuchungen nahe verwandter Arten. Die Kenntnis der in die Pheromonproduktion und -wahrnehmung involvierten Gene ermöglicht die Anwendung von Methoden der molekularen Phylogenetik, um zu verstehen, wie allgemeingültig diese Muster bei den Tausenden von anderen Mottenarten sind, deren fortwährender Reproduktionserfolg auf dem System ihrer Sexualpheromone beruht.